The Jaw of the Piltdown Man
A Reply to Mr. Gerrit S. Miller
W. P. Pycraft, F.Z.S., A.L.S., Memb. Roy. Anthrop. Instit.
Zoological Department, British Museum
Science Progress 1917
[389] Some time ago Mr. Gerrit S. Miller of the Smithsonian Institute, Washington, was requested by Dr. Ales Hrdlicka, of the United States National Museum, to compare a set of casts of the skull of the Piltdown man, supplied by the British Museum, with the skulls of anthropoid apes. This comparison was undertaken with the perfectly legitimate purpose of arriving at an independent opinion as to the conclusions arrived at by Dr. Smith Woodward, and other British anthropologists, as to the nature of the Piltdown remains. The result of that inquiry has been to convince Mr. Miller that the cranial portions of this now famous skull are unequivocally human, while the lower jaw is as certainly that of a chimpanzee. 1
In his summary
2 he tells us that "The Piltdown remains include parts of a brain-case showing fundamental characters not hitherto known except in members of the genus Homo, and a mandible, two molars, and an upper canine showing equally diagnostic features hitherto unknown, except in members of the genus Pan. 3 On the evidence furnished by these characters the fossils must be supposed to represent either a single individual belonging to an otherwise unknown extinct genus (Eoanthropus ) or to two individuals belonging to two now-[390]existing families (Hominidæ and Pongidæ).... In order to believe that all fragments came from a single individual, it is necessary to assume the existence of a primate differing from all other known members of the order by combining a brain-case and nasal bones possessing the exact characters of a genus belonging to one family, with a mandible, two lower molars, and an upper canine possessing the exact characters of a genus belonging to another. Thus must be associated in a single skull: (a ) one type of jaw with another type of glenoid region, (b ) one type of temporal muscle origin with another type of temporal muscle insertion, (c ) a high degree of basicranial adjustment to the upright position with absence of that corresponding modification in the lower jaw called for by all that is now actually known of the structure of the brain-case and mandible in primates, and (d ) a protruding lower jaw with a form of nasal bone not elsewhere known except in connection with a contracted upper dental arch. In each instance the opposed characters are sharply defined and easily recognisable in the fossils...."It is difficult to reconcile these very dogmatic statements, which have a semblance of statements of act with Mr. Miller's remarks in the opening passages of this summary: "The fossils are so fragmentary that their zoological meaning will probably remain a subject of controversy."
There does not seem to be much room for doubt about the "zoological meaning" of these remains; indeed Mr. Miller, in his main thesis, is very emphatic in every statement he makes thereon. He creates an impression indeed of having gone very thoroughly into the matter, with a wealth of material at his disposal enabling him to demolish completely the now generally accepted views as to the character of the skull of the Piltdown man. A very brief study of his arguments will show, however, that they are based on assumptions such as would never have been made had he not committed the initial mistake of overlooking the fact that these remains–which, by the way, he has never seen–are of extreme antiquity, and hence are to be measured by the standards of the palæontologist rather than of the anthropologist. This unfortunate lack of the right perspective has caused him to overlook some of the most significant features of these remains, and has absolutely warped his judgment in regard to the relative values [391] of the likenesses between these fragments and the skulls of the chimpanzee which he has so woefully misread.
When Mr. Miller commits himself to the statement, "Thus must be associated in a single skull: (a ) one type of jaw with another type of glenoid region," he implies that there exists a relationship between the mandible and the glenoid cavity which has no existence in fact. With the whole mammalian phylum to choose from he will seek in vain for data which will enable him to foretell, by an inspection of the glenoid cavity alone, what was the form of the jaw articulating therewith. What Mr. Miller appears to mean is, that because the glenoid cavity of the Piltdown man is of the type characteristic of modern man, therefore the mandible must be in keeping therewith, that is to say it must possess a "chin," and must be horse-shoe shaped. This does not in the least follow, and there is less reason to expect it in the present case because the cranium has not lost all its primitive characters, though he apparently assumes the contrary. What he really means becomes apparent when his remarks on p. 17 are compared with his indictment under heading "(c )," where he protests that it is impossible to associate in this skull "a high degree of basicranial adjustment to the upright position" with the "absence of that corresponding modification in the lower jaw called for by all that is now actually known of the structure of the brain case and mandible in primates." In the first place there is good reason for believing that the Piltdown man had not yet fully attained to the upright position, agreeing in this respect with the Heidelberg man. In the second the mandible does not present "the exact characters of a genus belonging to another family." The "absence of mechanical unity" between the mandible and the postulated face of this skull referred to on p. 17 is a purely imaginary absence. The jaw of the Piltdown skull, applied to a recent human skull of the same length as the Piltdown skull–of a Torres Straits Islander in the British Museum, to be precise–projects, at the incisors, no more than 3 mm.! Mr. Miller proposes to hang a lot on 3 mm. if he insists on his interpretation. As a matter of fact the Piltdown skull, even with this jaw, was less prognathous than in many modern men. This much is demonstrated by the restoration of the skull made in the American Museum of Natural History, [392] New York. For this has a facial angle of no more than 85o, which harmonises completely with the alveolar index of 105. In the British Museum are several skulls with facial angles ranging from 88o to 90o, and an alveolar index of as much as 110. Nor does it follow that because the toothrow in the Piltdown jaw was straight, as in chimpanzees, therefore , the rami must have run parallel with the sagittal plane. The relation to this plane is determined, not by the toothrow but by the intercondylar width. In megadont examples of modern man a straight toothrow frequently occurs, as I shall show later. His contention that the nasals, which he agrees to accord to the Piltdown skull, cannot possibly be associated with this jaw, is, like the rest of his argument, but a crude deduction founded on false premises. So much for his summary. Now for his details.
I will begin with the cranium. As to the human character of this there can be no two opinions. Mr. Miller himself is very emphatic on this point, building thereon much of his case against the human character of the jaw. He has missed, however, some of its most interesting features owing to his obsession in regard to the mandible. In the squamosal or "temporal," for example, the bone which he has designated as the type of Eoanthropus, he has failed to appreciate the fact that it is remarkable as much for its negative, as for its positive, characters. Having regard to its undoubted age one would have expected to find the simian spur on its anterior border which articulates, in all the apes, and in monkeys, and in many of the lower humans, with the frontal. We should also have expected to find a shallow glenoid cavity, as in the anthropoid apes, and many of the lower human types. Instead, this fossa is of most unusual depth, so much so that I have found but few modern skulls displaying a like depth. The eminentia articularis, on the other hand, is not only very extensive, but unusually oblique, which again is a peculiar character. In the form of the petrous portion it is most emphatically human, but here again it is peculiar, since it exceeds in length that of modern skulls by no less than 10 mm., giving a total breadth to the skull of no less than 20 mm. in excess of modern skulls. That Mr. Miller has entirely missed this feature is shown by a footnote (p. 17 of his memoir) in which he remarks that "...it is perhaps not safe to assume that the distance [393] from one glenoid cavity to the other was as great as in recent Homo...." As a matter of fact, as we have just pointed out, the skull of Eoanthropus must have been no less than 20 mm. wider than in modern Homo !
He is no less unfortunate in his interpretation of the temporal fossa, and in the inferences he draws from that interpretation. "The anterior border of the (temporal) muscle," he remarks, "appears to have extended upwards on to the frontal with somewhat unusual abruptness, an impression that may be heightened by the way in which the bone is broken." There is no room for doubt in this matter. The anterior border of the muscle did extend upwards with "unusual abruptness." The way in which the bone is broken has nothing whatever to do with the matter. A line drawn from the bregma to the external orbital process and compared with a similar line drawn across any modern human skull will at once demonstrate this point. But the abrupt trend of this portion of the temporalis fossa is very closely approached in many modern skulls of low races. What is unusual about this arc is due in part to the lowness of the cranial roof, and in part to the shortness of the frontal; which brings the coronal suture nearer to the upward limb of the arc in question. The backward extension of this muscle Mr. Miller has, I venture to think, underestimated, and he has failed also to notice the peculiar sinuous curve of its superior border. Furthermore, it is surely unfortunate to contrast the area of this muscle with that of the great apes in relation to the size of the animal ; such a comparison should surely be between the size of the head in the two groups. But in any case he is inaccurate as well as inopportune in his statements in regard to this muscle in the great apes. According to him, in Simia, or Pan as he prefers to call this genus, the temporal muscle meets its fellow in the middle line in adult individuals. This is true only of some males, belonging to some species, or sub-species. But even here the comparison lacks point. The muscle succeeds in reaching the middle line only because of the small size of the brain-case, but that factor he leaves entirely out of consideration.
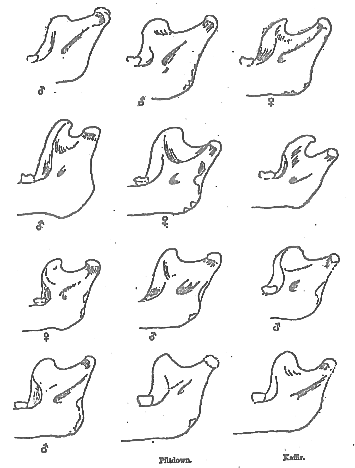
When he comes to treat of the area of insertion of this muscle his distortions of fact could not well go further. The form of the coronoid process and the ascending ramus of the [394] mandible, he insists, are so like what obtains in the chimpanzee that that likeness can be due only to one factor–the pull of a temporalis muscle such as obtains to-day in living chimpanzees, a muscle, he is at great pains to show, which differs profoundly from that of the human temporalis which covered the parieties of the skull of the Piltdown cranium. In his anxiety to prove this point, he proves too much, for if his main argument is sound, then the ascending ramus and coronoid process must present uniform characters in all chimpanzees. A very cursory survey of a number of jaws of these animals will suffice to demonstrate the baselessness of such an assumption. The form of the coronoid process, and of the ascending ramus, in regard to its width, is apparently governed by the size of the fossa bounded externally by the jugal bar and internally by the cranial wall. Where this fossa is long and wide, relatively to the length of the skull, the ascending ramus is wide, and the coronoid process is low. But the form of the sigmoid notch varies also, as a glance at the accompanying illustration (fig. 1) will show. It is clear, therefore, that Mr. Miller has placed a quite exaggerated importance on the likeness which seems to obtain between the form of the coronoid process and the sigmoid notch in the Piltdown skull and that of the two chimpanzees which he figures in his memoir. That likeness, by the way, is so close that I venture to suspect that it is largely due to that process of "mutilation" to which he tells us he has submitted these jaws in order that they made be made comparable to the Piltdown jaw. I have had the good fortune to examine far more jaws of chimpanzees than has fallen to the lot of Mr. Miller, and only in one instance have I come across even an approximate likeness. But the case is otherwise when a comparison is made with the jaws of prognathous human races, where a short and wide ascending ramus is common. But even if the likeness between these jaws is occasionally as close as Mr. Miller maintains, it adds but little to the strength of his arguments. In an investigation of this kind we have to take the sum of a large number of differences and resemblances; we cannot base far-reaching conclusions on trivial characters, such as he so often selects. Mr. Miller indeed has staked well-nigh everything on his interpretation of the coronoid process. This is unfortunate for his case, for
Fig. 1.– The ascending ramus of ten chimpanzee jaws, seen from the inner side, and compared with the same region in the Piltdown jaw, and that of a Kaffir. The extreme variability of the sigmoid notch, and of the size and shape of the coronoid process, show that no value whatever can be placed on inferences as to the size and shape of the temporalis muscles inserted here. The Piltdown jaw more nearly resembles that of the Kaffir than that of the chimpanzee.
[396] if the form of this process reflects so intimately the form, and mass, of the temporalis muscle that he can tell from its inspection in the Piltdown jaw what was the precise character of the temporalis inserted thereon, then the coronoid process of male chimpanzees' jaws in which the temporalis extended to the middle line of the skull should be readily distinguished from jaws in which this muscle failed to reach the median line, as in some males, and most female chimpanzee jaws. Yet such discrimination is impossible. On the contrary, the precise factors which mould the form of the coronoid process and of the sigmoid notch so far defy discovery, if only because of the extreme variety which they present. Typically, in the chimpanzee, the sigmoid notch, on which the form of the coronoid process largely depends, runs directly downwards and forwards, in a straight line from the condyle to the very base of the process in question, which commonly ends in a hook-shaped and backwardly directed summit. In some individuals, however, this notch is excessively deep and narrow, when the coronoid process closely approaches the condyle, as a glance at the accompanying figures will show. A notch of this latter type seems always to be associated with a very small jugal fossa. But save for this apparent correlation between the form of the jugal fossa and the ascending ramus of the mandible, no other governing factors in these extremely variable features seem discoverable. The dissimilarities of the ascending ramus of the mandible of the chimpanzee is remarkable, having regard to the restricted area of distribution of this animal. To find its parallel among human jaws all the races of the world must be sampled.
Great stress is laid on the significance and importance of the "secondary ridgelets" associated with the "ridge which extends upwards from the base of the coronoid process which gives extreme strength of attachment to the muscle fibre. This stage of roughening on the mandible is associated in the chimpanzee with the closest approach of the upper end of the muscle to the median line of the brain case and especially with the formation of a sagittal crest. It is well marked in the Piltdown jaw." These very circumstantial statements have no support in fact, at any rate in any of the specimens I have examined. The "ridgelets" are nowhere very obvious, and are no better developed in male than in female jaws, and [397] they are not present in the Piltdown jaw. The "highly developed strengthening ridge characteristic of the genus Pan " which runs downwards and forwards from the mandibular condyle to terminate above the dental foracen, though a common, is by no means a constant, feature of the chimpanzee jaw, and in some modern human jaws is almost as well marked as in the Piltdown jaw. Nor is the presence of the mylohyoid ridge a constant feature of the human jaw; in some, especially in low races, it may be barely perceptible. Below the missing condyle Mr. Miller insists the hinder border of the jaw "shows more lateral compression" than he has been able to find in any specimen of Homo. This failure can only be attributed to the limited number of human skulls to which he has access, an inference which also accounts for his apparent impression that the pterygoid fossa "immediately below the articular surface of the condyle is an invariable feature of the human jaw." It is not, and furthermore it is present in some chimpanzee jaws. Mr. Miller finds no indication of this fossa in the Piltdown jaw. Surely this is not a matter for surprise, having regard to the fact that the whole of the area in which this fossa is lodged is missing! Finally the form of the glenoid cavity of the squamosal or "temporal" bone affords but an indifferent guide to the form of the mandibular condyle, as a very cursory examination of a number of human crania will suffice to demonstrate. In some modern skulls this cavity is as deep as in the Piltdown skull, in others well nigh as shallow as in the chimpanzee, but there is no corresponding modification of the condyle.
Mr. Miller is always most emphatic where contradiction is most difficult, as for example when he elects to enlarge upon the character of missing elements in this jaw. Nowhere is this more apparent than in his comments on the missing condyle, but even here he is inconsistent. For on page 14 he remarks that "The articular process is worn off to the level where it begins to widen to form the base of the condyle." Yet, on the very next page (p. 15), he would have it believed that at least a portion of the condyle remains. This is the only interpretation possible on his statement, "Hence in order to fit its articulating surface to that of the skull it would be necessary to imagine an abrupt change of plan in the few millimetres of condyle that have been lost" (italics mine). What [398] does Mr. Miller mean by a "few millimetres"? Certainly less than I2, and probably 15 millimetres are missing. And this amount amply suffices to show that the support for the glenoid surface of the condyle, when restored by producing the parieties of the mandible upwards, takes the contour characteristic of the average human jaw much more readily than of the average chimpanzee jaw, which, by the way, commonly shows extremely prominent ridges and tubercles, for the insertion of the internal pterygoid muscle (fig. 2), though jaws in which these ridges are but slightly developed are also found. They are never so prominent in the human jaw, and they are barely perceptible in the Piltdown jaw.
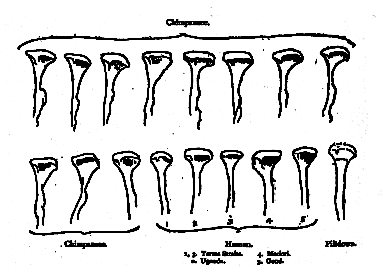
Fig. 2.–Outlines showing the contours of the posterior border of the ascending ramus, and of the condyle, in the jaws of chimpanzees and in human jaws. The condylar end of the sigmoid notch is just indicated in each case. It is clear that the Piltdown jaw more nearly conforms to the human than to the chimpanzee type.
In regard to the comparisons drawn between the jaws of chimpanzees and recent human jaws, and set forth in a Table of Measurements on p. 20 of Mr. Miller's memoir, there are several points which call for comment. Thus, in com[399]paring the length of the mandible at the alveolar level he gives 101.8 mm. as the maximum length in recent Homo. In the collections under my charge at the British Museum are jaws which, in this particular, range from 101 mm. to 115 mm., the latter the jaw of a Torres Straits Islander, that is to say only 5 mm less than the Piltdown jaw. The distance from the posterior border of the mandible to the front of M1 in recent Homo, according to Mr. Miller, attains its maximum at 75.6. In the British Museum collections we have recent human jaws ranging from this up to 88 mm., the highest being that of a Moriori. The maximum diameter of recent Homo of the ascending ramus at the alveolar level in this table is given as 46.8 mm. In the British Museum are jaws ranging up to 50 mm. In the chimpanzee, according to Mr. Miller, the maximum is 52 mm. Jaws which I have measured in the collection which Lord Rothschild has generously placed at my disposal range from this up to 55 mm. In Mr. Miller's table it will be noticed that his figures are all calculated to accentuate the differences between the Piltdown man and modern man, and the agreement with the chimpanzee. These revised figures lend no support to Mr. Miller; on the contrary, they place the Piltdown jaw well within the limits of human variation.
And now as to the teeth. "The two molars," we are told, "show no indication of the beginning of a curve in the toothrow. The main axis of the first tooth is continued by that of the second in a line passing as far to (the) inner side of (the) condyle as in the Pongidæ. In front of the first molar the entire hinder border of the alveolus of pm2 is plainly visible. It shows that the missing tooth was fully as large as in the great apes, and that the toothrow did not become sharply weakened at the point where this conspicuous change takes place in all known Hominidæ." These statements are to be read with his earlier arguments on p. 5. Here he tells us that "The toothrow in the Hominidæ is narrowed and weakened in front of the molars, the change taking place abruptly with (the) posterior premolar. Each premolar is single rooted, and the crown area is less than half that of the first molar.... Among the great apes the robust character of the toothrow is carried forward through the large, double-rooted premolars to the strongly functional canine.... A line joining the middle [400] of (the) posterior of m2, with the middle of (the) interior border of m1, will if continued forward in front of (the) incisors, converge rapidly with the sagittal line similarly extended. In the great apes ... a line passing through the middle of (the) posterior border of m2 and the middle of (the) anterior of m1 is essentially parallel to the sagittal line. In the Hominidæ the inward curve of the toothrow normally begins with the first lower molar. The axis of this tooth prolonged backwards diverges rapidly from a line parallel to the sagittal plane and crosses the posterior of m2 on (the) outer side of (the) middle; continued still further it passes through the condyle. That of the second tooth similarly prolonged, while diverging slightly from a line parallel to the sagittal plane, passes considerably to (the) inner side of (the) condyle. In all living genera of great apes, and in the fossil Propliothecus, Dryopithecus, and Sivapithecus, the axes of the two teeth lie in one line essentially parallel to the sagittal line, and passing further to (the) inner side of (the) condyle than is the case with the axis of m2 in the Hominidæ." The reference to the sagittal plane must be ignored since, according to Mr. Miller, the Piltdown jaw is that of an ape; regarded as a human jaw the greater intercondylar width of necessity causes a wide divergence from the median plane of the condyles.
The foundations of this argument are very insecurely laid. To begin with, there is no justification for the assumption which Mr. Miller makes, that the toothrow in chimpanzees is invariably straight. It is indeed commonly so, but an examination of a sufficiently large series of jaws will show that in some the molars are ranged along a backward and inward curve, and such instances I have reason to suspect are indicative of racial differences. But accepting Mr. Miller's interpretation, the application thereof fails, if only because the toothrow in human megadont jaws is also, commonly, as straight as in chimpanzees. Nor is there any justification for the statement that in the chimpanzee jaw the "robust character of the toothrow is carried forward through the large double-rooted premolar to the strongly functional canine," while in "all know Hominidæ" the toothrow becomes "abruptly weakened" by the conspicuously inferior size of the premolars; for in some chimpanzee jaws the premolars are emphatically small, while in megadont human jaws they may be of large size.
[401] As to the statements in regard to the axes of the molars, in relation to the condyle, they are not only inexact in regard to the chimpanzee, but they are equally inapplicable to the human jaw; since in megadont jaws the axis of m1 passes directly backwards, and is coincident with the axes of m2 and m3, as in the cases where this applies to the chimpanzee. Further, the common axis of the molars of the chimpanzee does not invariably, if produced backwards, pass further to the inner side of the condyle than in the Hominidæ. On the contrary, in some chimpanzee jaws this axis takes the same course as in the human jaw, that is to say, it touches the condyle. Mr. Miller's statements in regard to these relationships thus show either a very superficial acquaintance with the morphology of the human jaw, or a habit of basing far-reaching conclusions on ill-digested data.
Not the least characteristic feature of the teeth of the Piltdown jaw is their state of wear, the cusps having been worn down so as to reduce the crown to a perfectly level surface. Mr. Miller tells us that he has found teeth in the jaw of a chimpanzee worn in a precisely similar manner, and he gives a photograph purporting to bear out this statement. Yet no impartial critic will agree that this photograph in the least supports his statement. Of all the teeth which I have examined, and these represent at least twice as many as Mr. Miller has examined, I have failed to find one which can in any way be compared with these teeth of the Piltdown jaw in this matter of wear. But be this as it may, the determination of the generic identity of this jaw does not turn upon the state of wear of the crowns of the teeth, but upon a number of characters of far greater importance. Nor does their size affect the question, though according to Mr. Miller this is the only character by which they can be distinguished from the teeth of the chimpanzee, which, he asserts, are never so large as in the Piltdown jaw. This is not so. I have recently found teeth in the jaw of a chimpanzee from the Belgian Congo which are quite as large as those of the Piltdown jaw.
The molars of the Piltdown man, materially reduced as they are by wear, are still much more hypsodont than any chimpanzee teeth which I have yet seen, and this is true even if these worn teeth are compared with unworn chimpanzee teeth, which, by the way, are not, as Mr. Miller seems to suppose, of [402] so uniform a character that observations made will apply to all chimpanzee jaws. As a matter of fact, variety in the details of the contour of the crowns, and in the size of the cusps, as well as in the actual size of the teeth, show as great a range as in human jaws. Commonly the long axis of the lower molars of the chimpanzee exceeds that of the transverse axis, as is also the case in some modern human yaws, but it is by no means unusual to find teeth wherein these axes are equal, as is the rule in modern human teeth
Even where the teeth of the chimpanzee are as large as those of the Piltdown jaw, and such cases are rare, there is no difficulty in distinguishing between the molars of the ape and those of the man. In the Piltdown jaw the protoconid, metaconid, and hypoconulid are conspicuously larger than in those of the largest chimpanzee tooth, and these differences are still more marked in the case of the normally smaller teeth characteristic of the chimpanzee. Furthermore, the sulci dividing the cusps one from another are longer, and far more conspicuously marked, than in any human teeth, including the teeth of the Piltdown jaw (fig. 3, B). The widest part of the crown, in the chimpanzee tooth, is immediately above the roots, the grinding surface being conspicuously less in diameter. In human teeth, including the teeth of the Piltdown jaw, the crown passes almost insensibly into the root, and is not perceptibly wider or longer at its base than at its grinding surface; the reverse is the case with the molars of the chimpanzee. It is idle indeed to pretend that the molars of the chimpanzee are indistinguishable from those of the Piltdown jaw. As Prof. Keith has already remarked, radiographs of the Piltdown jaw show that they are of the typical "taurodont" type, therein differing conspicuously from the molars not only of the chimpanzee but of all the great apes.
A careful study of the unworn chimpanzee molars carried on side by side with human molars, also unworn, will speedily show yet another important differences between them. Those of the chimpanzee differ, for example, emphatically from all human teeth in having the external cusps–protoconid, hypoconid, and hypoconulid–sharply defined by deep sulci (fig. 3, B), and this is true even where the hypoconulid is degenerate. The contours of the grinding surface at their circumference vary so widely that no importance can be placed thereon in this [403] connection. But all chimpanzee teeth agree in that the otoconid and metaconid are united by a broad ridge showing a steep face posteriorly, and a deeply grooved upper surface (fig. 3. B). This is a feature which never occurs in human teeth where the ridge slopes gradually to the bottom of a cruciform valley, while in chimpanzee teeth the centre of the crown is occupied by a cup-shaped depression. Traces of this depression are to be found in some megadont human jaws, apparently also in the Piltdown jaw. Traces of the groove at the top of the ridge connecting the protocone with the metacone occur commonly in human molars. The cruciform valley, characteristic of the human molar, cuts the crown into four subequal moieties: this is never the case with the chimpanzee molar; since when lines answering to these valleys are drawn across the tooth, the transverse crosses the longitudinal axis obliquely.
Not the least characteristic feature of the worn surface of the teeth is the extreme thinness of the enamel, wherein they differ most emphatically from the teeth of the Piltdown jaw and from modern human teeth. Yet, curiously enough, Mr. Miller brushes this point aside as of no importance. And no less characteristic is the condition of the grinding surface of the crowns of worn molars of the chimpanzee. Contrary to Mr. Miller, I have in no single instance yet found these teeth with the surface worn to a perfectly level table at right angles to the vertical axis of the tooth. This is a common feature of human teeth. and is most conspicuous in those of the molars of the Piltdown man. In those of the chimpanzee, on the contrary, the outer sides may be worn down to the very base of the enamel while the cusps of the inner side are reduced to no more than half their original height. That the molars of the chimpanzee should occasionally present a perfectly flat surface need occasion no surprise, since, on the other hand, human teeth occasionally present the obliquely worn surface characteristic of the Chimpanzee, as for example in the case of the Moriori and some ancient Egyptians. This being so, the extremely flat surfaces of the crowns of the Piltdown molars should have given Mr. Miller occasion for reflection, having regard to the rule which undoubtedly obtains among the chimpanzees in this particular. At least it should have occurred to him that the wear of the teeth [404] was no safe guide to the form of the glenoid cavity, since it is clear that a precisely similar result of wear may accrue whether the glenoid cavity is deep, as in the human skull, or shallow, as in the chimpanzee.
And now as to the canine. Mr. Miller's arguments to prove that Dr. Smith Woodward is entirely wrong in his view that this is the right lower canine are founded upon assumptions for which no warrant can be found on an appeal to fact. "In all living great apes," he remarks, "the postero-internal surface of the lower canine is convex." This is true only of the tooth in transverse section: in a longitudinal section it is concave. Nor is he more happy in his statement that "No matter how long a lower canine may have been in use it never assumes the form seen in Eoanthropus, nor does it lose all trace of the original convexity of its inner portion." Mr. Miller's experience of worn canines in the chimpanzee is evidently limited, or he would not have failed to find examples which flatly contradict him when he says that all traces of the original convexity of the inner surface of the tooth are never lost. How largely he has substituted "intuition" for investigation is shown by his assurance that "a mechanical interrelation of the teeth, such as would produce a worn surface" presenting a wide shallow concavity directly backwards and inwards, "is not only unknown among primates, but (in a lower canine) I have never been able to find any mammal with the upper and lower teeth so arranged that it could exist." This may very well be true, so far as Mr. Miller is concerned, but in the British Museum are two chimpanzee jaws (1.8.9.4. and 1.8.9.10.), in the collection of Lord Rothschild is a third, and in the Royal College of Surgeons' Museum is a fourth, achieving Mr. Miller's standard of the impossible. In the British Museum specimens the worn surfaces, however, present a slight twist, recalling that of a propeller, but in that in Lord Rothschild's museum the resemblance to the Piltdown tooth is extraordinarily close. In studying the worn surfaces of these teeth in the great apes the aspect which most impresses one is not the similarity, but the remarkable dissimilarity, they present, a phenomenon due to variations in their size, and the angle at which they stand in the socket, for these vary the incidence of wear by the opposing teeth. In some cases attrition is caused, not by the
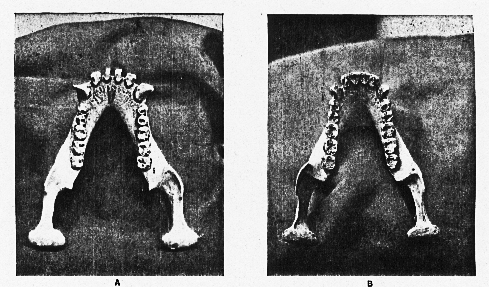
Fig. 3.–Jaws of Chimpanzee, showing the lateral torsion of the alveolar border, which carries the teeth obliquely across the body of the jaw. In A (No. 61.7.29.10 Brit. Mus. Coll.) the mesial aspect of the ramus, from the level of M2, is much inflated, forming a conspicuous lateral cushion meeting behind the incisors and overhanging a spacious "genial pit." In B the "lateral cushion" is indicated merely by a slight bulging of the inner wall of the mandible. The symphysis presents a steep face, thereby considerably reducing the size of the genial pit. The molars in this jaw are slightly larger than in the Piltdown jaw. This photograph is from a specimen belonging to the Belgian Government for the Congo.
[405] upper canine, but by the lateral incisor. Hence it is that in no two canines, even in the same jaw, are the results of wear the same. This being so, even if the Piltdown jaw were indubitably that of a chimpanzee, it would be unreasonable to expect to find its exact counterpart in modern chimpanzees. There is nothing in short. in the character of the worn surface of this tooth which is inconsistent with the position in the lower jaw which has been assigned to it.
That this canine is not that of a chimpanzee is surely abundantly proved by the fact that it lacks all traces of the "heel" at the postero-external angle of the base, as was pointed out by Dr. Smith Woodward, as well as of the "cingulum" which is present in the canine of all chimpanzees, in some examples of which it is extremely well developed.
While I am convinced of the correctness of Dr. Smith Woodward's interpretation as to the cause of the wear in the Piltdown tooth, the worn surface of the teeth in the jaw in Lord Rothschild's collection, already referred to, lends colour to the contention of Prof. Keith that this wear was caused, not by the upper canine, but by the upper lateral incisor. That, however, is a matter of detail of no great importance.
Mr. Miller may urge that my arguments do not affect his case in the least, since he associates this tooth with the upper, and not with the lower jaw. But there is, if possible, even less to support him in this view. Nevertheless Mr. Miller is not alone in this association, but it is almost incredible that it should have received serious consideration. Those who hold this opinion, indeed, do so either because they have not had an opportunity of studying the actual tooth, or from a wholly insufficient examination of the evidence afforded by the canine of the chimpanzee.
In the first place the root of the Piltdown tooth, both in its curvature and in its section, is quite unlike what is demanded of an upper canine, since it has a strong outward and only a very slight backward curvature, and presents a very conspicuous flattening throughout its entire internal aspect. In the second place the condition of its worn surface is quite inconsistent with a position in the upper jaw. In all the jaws of chimpanzees which I have examined the wear of the upper canine, where it is at all comparable to that of the Piltdown [406] tooth, ends abruptly, forming an overhanging ledge at the base of the tooth, whereas in the Piltdown tooth it presents a continuous curve. But, further, the worn surface of the Piltdown tooth looks backwards and outwards, a form of wear impossible in an upper canine, while it is demonstrably possible in a lower canine. The jaw in the College of Surgeons already referred to (No. 1, Du Chaillu Coll. Gaboon, Male) shows a lower canine which in this matter of wear is almost identical with that of the Piltdown tooth; it differs, indeed, only in that its posterior external edge is slightly worn apparently by the upper canine, the rest of the surface seeming to have been worn by the upper lateral incisor. The upper canines of this skull have been broken and worn to mere stumps, yet showing a form closely resembling the upper canine postulated by Dr. Smith Woodward in his restoration of the: Piltdown skull.
Two points of paramount importance in the conformation of the Piltdown jaw have been entirely overlooked by Mr. Miller. The first of these concerns the conformation of the inner surface of the body of the jaw. In chimpanzees this shows very striking differences, forming, when extremes are compared, two well-marked types (fig. 3). In the one (fig. 3, A) the inter-ramal area, from the symphysis backwards as far as the level of M2, has a curiously inflated appearance, so that the teeth seem to arise from a cushion-like bed, which, anteriorly, overhangs the genial pit. In the other this inner wall may dip downwards from the teeth almost as abruptly as in human jaws (fig. 3, B). But even here the differences between the human and the chimpanzee jaw are readily apparent, since in the chimpanzee this inner wall shelves downwards, and inwards, towards the symphysis, which is always much deeper, and never so steep, as the human jaw. As to the actual symphysis in the Piltdown jaw no positive statements can be made, but there can be no possibility of doubt about the human character of the whole region in question which has been preserved. Mr. Miller's photographs of the inner aspects of the jaws, which he uses to demonstrate the community of descent between the owner of the Piltdown jaw and modern chimpanzees, succeed only in demolishing the theory which he has been at such pains to elaborate. For one can see at a glance, by the high lights, [407] which are the jaws of chimpanzees and which is the human –Piltdown–jaw. Between the two extremes seen in the jaws of chimpanzees every gradation will be found, but in no case would there be any possibility of confusing the Piltdown fragment, or any similar fragment of a modern human jaw, with similar fragments of chimpanzee jaws. This character alone suffices to demolish the whole of Mr. Miller's arguments.
But there is yet another test as to the human character of the Piltdown jaw which I venture to think is the most convincing of all. Mr. Miller makes much apparent capital out of the fact that when the jaw of a chimpanzee is compared with that of a modern man it will be found that the toothrow from the canines backwards, in the chimpanzee, are parallel, while in man the molars along the two sides of the jaw diverge; and he supports his statements by means of diagrams. No one doubts the correctness of these; the facts which he points out have long been familiar to anthropologists. But this comparison is absolutely valueless in the present connection, since we have but one ramus of the Piltdown jaw. This alone, however, furnishes two extremely important characters. One of these has already been discussed. The other may be briefly stated as follows.
If, in the jaw of a chimpanzee, a line be drawn down the middle of the toothrow from the canine backwards, and another be drawn through the ascending ramus entering by the posterior border and passing out through the anterior border, it will be found that the two lines converge in front of the canine (fig. 4). If these lines be drawn along the toothrow, and through the ascending ramus, of the jaw of a modern man they will be found to converge at a variable distance behind the articular condyle of the jaw; they may in rare cases run parallel, but in no case have I yet found them converging in front of the canine as in the chimpanzee. 4 The Piltdown jaw agrees with that of modern man (fig. 4).
The interpretation of these differences is not difficult. In the chimpanzee the toothrows of the right and left rami en[407]close a long, narrow, lingual space, so narrow that the ascending rami have to undergo a wide lateral extension in order to provide the intercondylar width necessary for the articulation of the jaw with the cranium. In the human jaw the teeth are arranged horse-shoe fashion, diverging rapidly posteriorly, hence the ascending rami have a relatively smaller lateral extension in regard to the toothrow. The fact, then, that the Piltdown jaw in this regard agrees with the modern human jaw suffices to show that in the arrangement of its teeth, and in the form of the enclosed lingual space, it differed as widely from that of the chimpanzee as does the man of to-day.
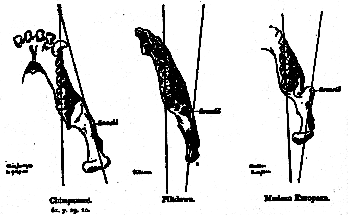
Fig. 4.–Lines drawn along the molars, and through the ascending ramus, to show that in the chimpanzee they converge in front of the canine, while in the human jaw they converge at a variable distance behind the condyle. The human character of the Piltdown jaw, judged by this test, is unmistakable.
That the Piltdown jaw does present many points of striking resemblance to that of a chimpanzee is beyond dispute. Dr. Smith Woodward pointed out these resemblances long ago, in his original description of the jaw. But Mr. Miller contends that because of these resemblances it is the jaw of a chimpanzee. It will be obvious, to those who will take the trouble to analyse the evidence wherewith he supports his arguments, that he has endeavoured, throughout, to confirm [409] a preconceived theory; a course of action which has unfortunately warped his judgment and sense of proportion. He has laboured under the further disadvantage of never having seen the original remains. This implies no reflection upon the accuracy of the casts supplied him from the British Museum, which were indeed most faithful reproductions, but it is humanly impossible to impart to plaster of Paris the subtle qualities of mineralized bone and enamel which count for so much in a task such as Mr. Miller set himself.
In order that I might do Mr. Miller full justice I have made a very careful study of the skulls of chimpanzees in the British Museum and the Royal College of Surgeons. These were supplemented by the fine collection in the museum of Lord Rothschild, which he generously placed at my disposal, sending them up to me at the British Museum to facilitate comparison. I therefore desire here to express my grateful thanks both to Lord Rothschild and to Prof. Keith for the very material help they afforded me.
Finally, I have submitted my objections to Mr. Miller's contentions to Dr. Smith Woodward, Prof. Elliot Smith, Prof. Arthur Keith, Prof. A. S. Underwood, and Dr. Broom, and with one accord they permit me to say that they are in entire agreement with me.
_____________________________________________________________________________
1 Since this was written Dr. W. K. Gregory, of the American Museum of Natural History, has published a memoir–Studies on the Evolution of the Primates–in which he throws the weight of his authority on the side of Mr. Miller; but since he accepts Mr. Miller's arguments without investigating the matter for himself, the objections herein set forth to Mr. Miller's contentions apply equally to Dr. Gregory.
2 "The Jaw of the Piltdown Man," Smithsonian Miscellaneous Collections, vol. 65, No. 12.
3 When Mr. Miller speaks of the genus Pan he means the genus Simia.
4 The line through the ascending ramus can best be taken by means of a "straight edge" held immediately above the sigmoid notch, but not necessarily over the tip of the coronoid process, which, both in man and apes, may be deflected outwards. At the same time another "straight edge" should be held over the toothrow.